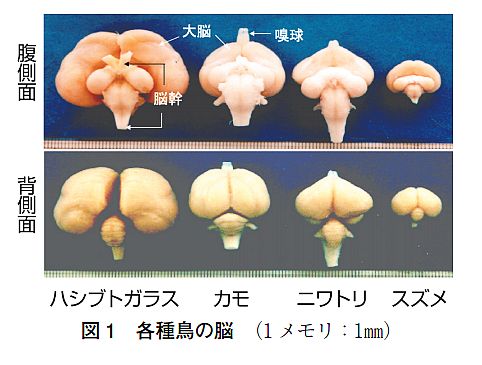
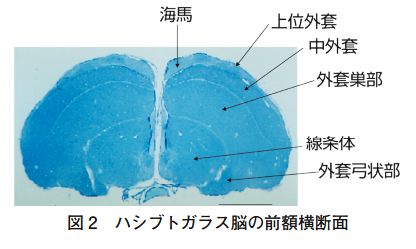
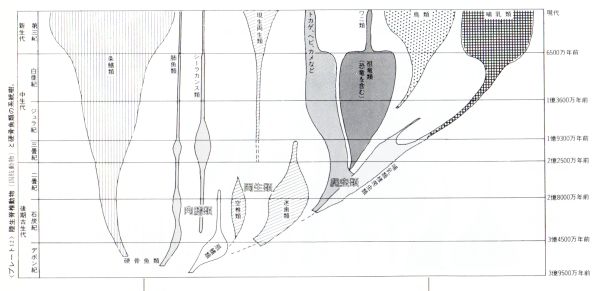
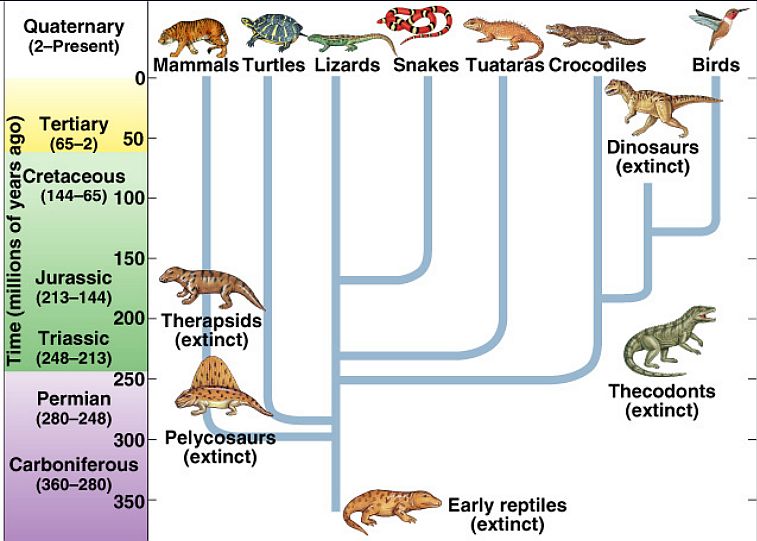
大脳の起源について一通り勉強してきた。
最近の遺伝子研究によって大脳の起源についての記述は大きく書き換えられつつある。
ただしそれはルーツ探しの旅のようなもので、Y染色体やミトコンドリアDNAのように、現存人類のところから遡っていく旅のようである。
そうやって我々はミトコンドリア・イブやY染色体アダムのところまで行き着く。
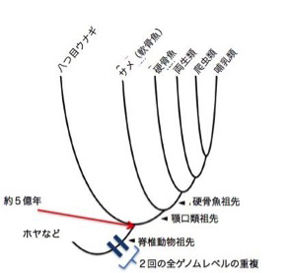
それは予想をはるかに越えて、5億年の昔、ナメクジウオを生み出した祖先までたどる旅となる。
ただ、そこで証明されるのは、ナメクジウオがすでに人類の脳を生み出す遺伝的能力を備えていたという事実であって、それが何故に発現してきたかの過程とは異なる。ルーツ探しの旅を決定論的に読み替えるのは有害無益である。
その上で、系統発生的には次のような大脳形成過程が想定できるだろうと思う。
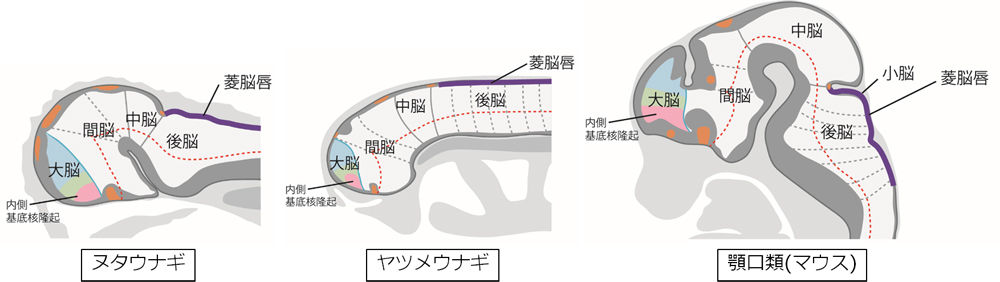
① ナメクジウオのレベルでの三脳構造の確立。② ヤツメウナギのレベルでの前脳前方への外套の形成③ 魚類(顎口類)のレベルでの外套の翻転と終脳の形成
ここで脳の分節構造を信じるならば、「外套→終脳」は、前脳が間脳と終脳に割れたのではなく、もともと前脳の前方に“もう一つの分節”の萌芽として内在した終脳原基が発現したものとして捉えるべきだろうということだ。
言葉で表現するなら、終脳は前脳より前方の最先脳(最終脳)であり、間脳は、結果として前から2番目の脳になったから間脳だということになる。
前脳が視床と視床下部という背腹(上下)2階建てになったのと同じく、終脳も大脳皮質(外套)と大脳基底核(腹側外套と外套下部)の2階建てになったのであろう。